 Văn phòng Học viện
Văn phòng Học viện
Khi nào và làm thế nào để tiêu diệt tế bào thực vật: Chiến lược lây nhiễm của nấm gây bệnh thực vật
1. Giới thiệu
Rất nhiều nghiên cứu khoa học đã phát hiện các chủng vi sinh vật, bao gồm cả vi khuẩn và xạ khuẩn, có khả năng đối kháng với các chủng nấm gây bệnh trên thực vật, cũng như là cơ chế đối kháng của chúng với những chủng nấm gây bệnh. Nhưng có bao giờ bạn đặt ra câu hỏi nguyên nhân tại sao các chủng nấm tấn công thực vật và cơ chế tấn công của chúng là gì? bài viết này sẽ làm sáng tỏ những câu hỏi đó. Nấm gây bệnh hại thực vật đã phát triển các kiểu sống và phương thức tương tác khác nhau với cây chủ của chúng. Một số loại nấm gây bệnh tổng hợp và tiết ra các hợp chất thứ cấp có độc ở lần xâm chiếm đầu tiên, tiêu diệt tế bào vật chủ và sống nhờ các hợp chất hữu cơ của tế bào chủ. Ngược lại, các loại nấm khác sống nhờ chất dinh dưỡng do vật chủ cung cấp trong thời gian dài và không tạo ra độc tố. Những chế độ dinh dưỡng tương phản này được gọi là hoại dưỡng (necrotrophy) và sinh dưỡng (biotrophy). Tuy nhiên, cũng tồn tại sự kết hợp của hai kiểu sống và chiến lược dinh dưỡng này, các loại nấm gây bệnh biểu hiện kiểu sống sinh dưỡng tạm thời khi bắt đầu xâm chiếm tế bào chủ sau đó là hoại dưỡng, kiểu sống này được gọi là bán sinh dưỡng (hemibiotrophs).
2. Sự hình thành cấu trúc lây nhiễm của nấm, sự xâm nhiễm vào cây và chiến lược dinh dưỡng
2.1. Nấm sinh dưỡng
Các nấm gây bệnh có kiểu sống sinh dưỡng bắt buộc như nấm gây bệnh phấn trắng và gỉ sắt phụ thuộc vào tính toàn vẹn của tế bào chủ mà chúng ký sinh. Để xâm nhập qua thành tế bào thực vật, các cấu trúc lây nhiễm chuyên biệt cao được hình thành (Hình 1A). Trong trường hợp nấm gây bệnh phấn trắng ngoại ký sinh, các ống mầm và giác bám sơ cấp được hình thành trên lớp biểu bì thực vật, sau này làm phát sinh các tế bào lây nhiễm gọi là giác bám (appressoria). Những tế bào này cho phép nấm gây bệnh xâm nhập vào thành tế bào bằng cách sử dụng sự kết hợp giữa lực và các enzyme phân hủy thành tế bào. Vật chất của các túi được vận chuyển bởi nấm sinh dưỡng bắt buộc tại vị trí xâm nhiễm có thể không chỉ là các enzyme phân hủy thành tế bào, mà còn là các phân tử tác động được tiết vào tế bào chủ để ngăn chặn việc kích hoạt các phản ứng phòng vệ. Sự hình thành và tiết các enzyme khác nhau phân hủy pectin, cellulose và protein xảy ra khi hình thành các cấu trúc xâm nhiễm chuyên biệt. Điều thú vị là trong quá trình sinh dưỡng bắt buộc, việc tiết ra một số enzyme phân hủy thành tế bào không dẫn đến sự hoại tử mô, có thể do thực tế là các enzyme phá hủy này có thể bị cố định bởi sự tương tác ion với các chất trao đổi ở thành tế bào thực vật và do đó hoạt động của chúng vẫn bị hạn chế ở các khu vực cụ thể (Horbach & cs., 2011).
Nấm sinh dưỡng bắt buộc làm lõm màng sinh chất sau khi xuyên qua thành tế bào thực vật, và màng sinh chất bao chặt chẽ quanh tế bào dinh dưỡng mới hình thành gọi là giác hút (haustorium) trong suốt vòng đời của nó. Enzyme ATPase có trong màng giác hút sẽ axit hóa không gian bên ngoài giác hút, do đó tạo ra sự chênh lệch pH (hay gradient pH) thúc đẩy quá trình đồng vận chuyển (symport) đường và axit amin-H+ được xúc tác bởi các chất vận chuyển phân bố trong màng giác hút. Giác hút không chỉ cần thiết cho việc hấp thụ chất dinh dưỡng mà còn được cho là nơi thuận lợi để tiết protein tác động (effector). Tuy nhiên, các protein tác động vẫn chưa được mô tả chức năng.
|
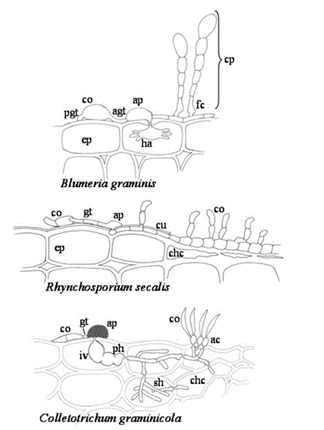
Hình 1. Cấu trúc lây nhiễm của nấm với kiểu sống khác nhau
(A) Nấm sinh dưỡng bắt buộc B. graminis hình thành ống mầm sơ cấp và ống mầm giác bám (pgt; agt), và giác bám (ap), xâm nhập vào tế bào biểu bì của vật chủ (ep) để tạo thành giác hút (ha). Sợi nấm phát triển trên biểu bì thực vật tạo thành bào tử ngủ (cp), tế bào chân đặc trưng (fc). (B) Bào tử đính chứa hai tế bào (co) của nấm hoại dưỡng R. secalis tạo thành ống mầm (gt) và giác bám (ap) trên bề mặt cây chủ của nó. Sau khi xuyên qua lớp biểu bì (cu), sợi nấm mỏng với các vách ngăn cách xa nhau phát triển dưới lớp biểu bì, tiếp theo là sợi nấm dày với các tế bào ngắn, mọc trên bề mặt lá và hình thành bào tử mới. (C) Sau khi nảy mầm của sự hình thành bào tử đính (co) và ống mầm (gt), nấm bán sinh dưỡng C. graminicola phân hoá một vòi bám bị hắc tố (ap). Bên trong tế bào vật chủ bị xâm chiếm đầu tiên, một túi nấm sinh dưỡng lây nhiễm (iv) và sợi nấm sơ cấp (ph) phát triển. Sau đó, sợi nấm thứ cấp hoại dưỡng, phát triển nhanh (sh) xâm chiếm lá và tạo ra đĩa đài (acervuli) (ac), nơi hình thành bào tử mới.
2.2. Nấm hoại dưỡng
Trong hầu hết các trường hợp, chiến lược lây nhiễm của nấm hoại dưỡng ít phức tạp hơn so với nấm sinh dưỡng bắt buộc. Đĩa bám được hình thành bởi các loài nấm hoại dưỡng điển hình như Alternaria, Botrytis, Cercospora, Fusarium, Helminthosporium, Ramularia, Rhynchosporium, Sclerotinia hoặc các loài Verticillium không dễ thấy và các sợi nấm xâm nhiễm được hình thành bên trong vật chủ khá đồng nhất. Việc tiêu diệt tế bào chủ được thực hiện bằng các chiến lược khác nhau như tiết ra chất có khối lượng phân tử thấp hoặc peptide độc tố hoặc bằng cách gây chết tế bào ở vật chủ bằng cách tiết ra các phân loại oxy hoạt hoá (reactive oxygen species / ROS).
Ở một số nấm hoại dưỡng gây bệnh, độc lực tương quan với khả năng tổng hợp độc tố. Việc giải phóng độc tố và/hoặc các phân loại oxy hoạt hoá dẫn đến phá hủy màng ngay lập tức, cung cấp chất dinh dưỡng cho nấm và quan trọng nhất là ngăn chặn việc kích hoạt các phản ứng bảo vệ của thực vật chủ.
Nấm hoại dưỡng không chỉ tiêu diệt vật chủ bằng cách tiết ra độc tố hoặc ROS, mà còn tiết ra một lượng lớn enzyme phân hủy thành tế bào để phân huỷ các polymer cấu tạo thành tế bào và thường gây tổn thương mô đáng kể. Việc phân huỷ thành tế bào có thể phục vụ hai mục đích, hỗ trợ sự thâm nhập và cung cấp nguồn carbohydrate. Do số lượng lớn các gen mã hóa các enzyme phân hủy thành tế bào có trong bộ gene của nấm hoại dưỡng gây bệnh thực vật, có thể hình dung rằng việc gây mất gen mục tiêu không làm giảm độc lực ở các thể đột biến. Tuy các enzyme phân hủy thành tế bào và các mảnh vỡ do sự phân hủy có thể tạo ra các phản ứng phòng vệ, nhưng thực tế là các gene mã hoá enzyme vẫn được duy trì hoặc thậm chí được nhân đôi đã củng cố mạnh mẽ lập luận rằng các enzyme phân hủy thành tế bào là không thể thiếu đối với độc lực của nấm gây bệnh thực vật.
Ở hầu hết các cây bị các bệnh do nấm hoại dưỡng tấn công, có thể quan sát thấy sự hoại tử của tế bào vật chủ ngay sau khi nấm xâm nhập. Các ống mầm phát triển từ bào tử ngủ của nấm tạo thành các vòi bám không rõ ràng (Hình 1B và 2A), xâm nhập vào lớp biểu bì của lá cây chủ, thường ở rãnh phía trên thành nếp lồi (anticlinal walls) giữa các tế bào biểu bì liền kề. Sau đó, nấm phát triển dưới biểu bì ở vùng giàu pectin của thành tế bào biểu bì bên ngoài. Các sợi nấm hình thành các khuẩn lạc non hầu hết mỏng và có vách ngăn rộng. Sau đó, hình thái sợi nấm thay đổi thành các cấu trúc có vách ngăn ngắn dày hơn tạo thành lớp đệm dưới biểu bì. Sự hình thành bào tử mới xảy ra bên ngoài lá ở phần mở rộng của sợi nấm, nhô ra khỏi chất nền (Hình 1B, 2A và B). Thông thường, không có triệu chứng bệnh nhìn thấy bằng mắt thường nào xuất hiện trong một thời gian dài sau khi xâm nhiễm. Tuy nhiên, sự hoại tử của các tế bào biểu bì (Hình 1 B) bắt đầu sớm trong quá trình ủ bệnh, do việc tiết ra các peptide gây hoại tử (necrosis-inducing peptides / NIP) (xem bên dưới), và tiếp tục cho đến khi mô đỡ của nấm thay thế các phần lớn của lớp biểu bì (Hình 2 B). Tại thời điểm này, các tế bào thịt lá cũng bắt đầu hoại tử, dẫn đến xuất hiện các triệu chứng bệnh rõ ràng. Chỉ ở những giai đoạn muộn này, sợi nấm đơn lẻ mới có thể được tìm thấy trong mô thịt lá bị phá hủy.
2.3. Nấm bán sinh dưỡng
Các nghiên cứu gần đây cho thấy tế bào chủ sau khi bị lây nhiễm bởi nấm gây bệnh hại cây không bị tiêu diệt ngay lập tức mà vẫn tồn tại trong giai đoạn lây nhiễm ban đầu. Các loài nấm này được gọi là nấm bán sinh dưỡng.
Người ta có thể suy đoán rằng dạng sống (kiểu sống) sinh dưỡng đã được hình thành như kiểu sống nguyên thuỷ của nấm gây bệnh và kiểu sống hoại dưỡng đại diện cho thành tựu tiến hóa gần đây hơn, kiểu sống bán sinh dưỡng phản ánh sự chuyển đổi giữa các kiểu dinh dưỡng này. Việc tiêu diệt tế bào chủ và chuyển sang kiểu sống hoại dưỡng thường xảy ra vài ngày sau khi thiết lập giai đoạn sinh dưỡng của nấm gây bệnh trong tế bào thực vật. Các giác bám định hướng lại hướng phát triển của sợi nấm: Trong khi ống mầm phát triển trên lớp biểu bì thực vật, sợi nấm xâm nhiễm bắt nguồn từ đáy của giác bám sẽ xâm lấn và xâm nhiễm vào mô của cây chủ. Thành của sợi nấm xâm nhập có những đặc điểm khác với các đặc điểm của ống mầm.
Các enzyme chitinase nội bào phân cắt chitosan kém hiệu quả hơn so với chitin, dẫn đến kết quả các sợi nấm xâm lấn được bảo vệ trong các giai đoạn sinh dưỡng của sự tương tác và làm giảm việc tạo ra các đoạn chitin có hoạt tính elicitor.
Một số đặc trưng, ví dụ: những thay đổi về đặc tính bề mặt của sợi nấm lây nhiễm, duy trì tính toàn vẹn của màng sinh chất của vật chủ và sắp xếp lại quá trình trao đổi chất của thực vật được biểu thị bằng sự hình thành các đảo xanh (green islands), hỗ trợ cho ý tưởng rằng nấm bán sinh dưỡng ở các thời điểm đầu của kiểu sống sinh dưỡng có thể được so sánh trực tiếp với nấm sinh dưỡng bắt buộc, nhưng các lập luận về chức năng thì còn hạn chế.
H2O2 do nấm tạo ra có thể là công cụ tiêu diệt tế bào chủ, như thường thấy trong các tương tác liên quan đến nấm hoại dưỡng. Đáng chú ý, nếu trong giai đoạn đầu sinh trưởng của nấm sinh dưỡng, tế bào chủ tạo ra H2O2 dẫn đến tế bào chết nhanh chóng, sự phát triển của nấm bị chậm lại. Sự tích lũy H2O2 được biết là đại diện cho một phản ứng phòng vệ có hiệu quả cao chống lại các bệnh gây ra bởi nấm sinh dưỡng, nhưng không chống lại các nấm hoại dưỡng. Điều này, cùng với những điểm tương đồng giữa nấm sinh dưỡng và nấm bán sinh dưỡng đã thảo luận ở trên cho thấy rằng sợi nấm sinh dưỡng sơ cấp ở nấm bán sinh dưỡng thực sự giống với giác hút của nấm sinh dưỡng. Do đó, nấm bánh sinh dưỡng có thể cho phép nghiên cứu nấm sinh dưỡng ở cấp độ phân tử, tương phản với nấm sinh dưỡng bắt buộc, những loại nấm này có thể tiếp cận được bằng các công cụ phân tử. Tuy nhiên ở thời điểm công bố bài báo này, tín hiệu trung gian chuyển đổi giữa nấm sinh dưỡng - nấm hoại dưỡng ở nấm bán sinh dưỡng vẫn chưa được tìm hiểu rõ.
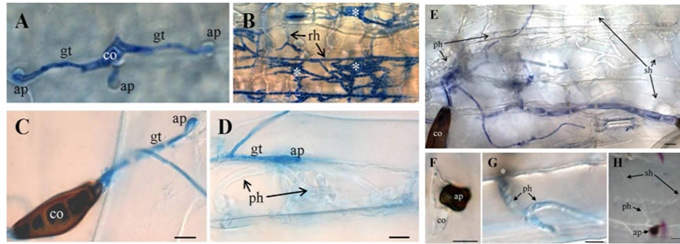 |
Hình 2. Giai đoạn đầu hình thành khuẩn lạc trên lá của nấm có kiểu sống khác nhau
(A và B) R. secalis. Trên lớp biểu bì của vật chủ (A), bào tử đính (co) nảy mầm với một ống mầm (gt) để tạo thành vòi bám (ap). Ở giai đoạn sau (B) t lá bị hình thành khuẩn lạc các sợi nấm chạy (rh), và các sợi nấm liên kết được gọi là stroma (*) được hình thành, nơi bào tử định mới được tạo ra. (C–E) B. sorokiniana bào tử đính (co) đa bào hình thành các ống mầm (gt) và vòi bám (ap) (C), và xâm chiếm lá để phân hoá sợi nấm sinh dưỡng sơ cấp dày (ph) (D). Ở giai đoạn sau (E), sợi nấm sơ cấp phát triển thành sợi nấm hoại dưỡng thứ cấp (sh), tiêu diệt tế bào chủ một cách hiệu quả. (F–H) C. graminicola. Tế bào đính (co) của nấm bán sinh dưỡng tạo thành một vòi bám có hắc tố (ap) trên lớp biểu bì của vật chủ (F), xuyên qua lớp biểu bì và thành tế bào và hình thành sợi nấm sơ cấp (ph) trong tế bào chủ (G). Vòi bám (G) nằm trong một mặt phẳng tiêu điểm khác (*). Sợi nấm thứ cấp (H) (sh) đại diện cho kiểu sống hoại sinh, xâm nhiễm mô chủ một cách nhanh chóng và gây chết tế bào chủ. thanh = 10 µm
Bài viết này đã chứng minh rằng nấm gây bệnh thực vật đã phát triển một loạt các chiến lược để lây nhiễm sang cây chủ, như được chỉ ra ở cấp độ hình thái bằng cách hình thành các cấu trúc lây nhiễm đa dạng, phương thức hấp thu dinh dưỡng và tiêu diệt cây chủ. Các chiến lược dựa trên ba kiểu sống của nấm kí sinh đó là sinh dưỡng, hoại dưỡng và bán sinh dưỡng. Nấm sinh dưỡng không làm chết mà chỉ hút chất dinh dưỡng từ cây chủ trong khi đó nấm hoại dưỡng gây chết tế bào và hấp thu dinh dưỡng từ cây chủ đã bị phân huỷ. Bên cạnh đó, kiểu sống bán sinh dưỡng là sự kết hợp của hai kiểu sống đã đề cập phía trên, nấm kí sinh chuyển đổi từ kiểu sống sinh dưỡng sang hoại dưỡng vài ngày sau khi xâm nhiễm vào cây kí chủ.
Theo Horbach, R., Navarro-Quesada, A. R., Knogge, W., & Deising, H. B. (2011). When and how to kill a plant cell: Infection strategies of plant pathogenic fungi. In Journal of Plant Physiology (Vol. 168, Issue 1, pp. 51–62). https://doi.org/10.1016/j.jplph.2010.06.014
Ý kiến bạn đọc




- Đang truy cập6
- Hôm nay1,797
- Tháng hiện tại6,079
- Tổng lượt truy cập2,385,842